
Le sang des Mammifères permet les échanges de matière et d’énergie entre les cellules. Le sang circule pour cela dans l’ensemble de l’organisme, dans les vaisseaux sanguins. La circulation du sang requiert pour cela un système moteur. C’est ainsi que le cœur permet la circulation sanguine, en se comportant comme une pompe. Le sang circule sous pression dans les vaisseaux sanguins : le cœur se contracte et la hausse de la pression entraîne le liquide. C’est pourquoi on parle de pompe cardiaque.
Comme tout organe, le cœur est composé de tissus avec leurs propres propriétés, des propriétés mécaniques, d’échange, et autres et qui en assurent le bon fonctionnement. Il va donc falloir se pencher un peu sur les tissus et les cellules du cœur pour en comprendre le fonctionnement. Et enfin, comme tout organe, son fonctionnement doit pouvoir être modulé par l’organisme pour répondre à ses besoins.
Ainsi, cet article se propose de discuter un peu de ces points. Il ne sera donc pas complet, mais s’essaiera à faire une petite introduction à la physiologie cardiaque. Les pré-requis ne sont a priori pas trop élevés ; vous ne devez être familier qu’avec un peu d’anatomie humaine et quelques éléments de structure cellulaire pour comprendre pleinement l’article.
- Avant-propos : généralités de physiologie
- L'anatomie cardiaque
- Les cellules cardiaques
- Le cycle cardiaque
- Le devenir du sang et les propriétés d'hémodynamique
- Régulation cardiaque
Avant-propos : généralités de physiologie
Pour comprendre pleinement le rôle du cœur, il est nécessaire d’aborder le concept de milieu intérieur de d’homéostasie du milieu intérieur.
Le concept de milieu intérieur a été développé par Claude Bernard ; les cellules de tout organisme échangent de la matière et de l’énergie avec leur milieu environnant. Pour des organismes unicellulaires comme des bactéries, il s’agit de l’environnement direct. Les échanges sont impactés par la composition de l’environnement : salinité, richesse en sucres et acides aminés, mais aussi chaleur.
Pour les organismes pluricellulaires comme les Mammifères, les cellules ne sont plus en contact avec l’environnement direct : les cellules internes échangent toujours, mais avec un milieu dont la composition n’est pas celle de l’environnement direct. Un peu comme, pour les cavernes, l’atmosphère est plus humide et plus fraîche alors qu’elles sont tout de même reliées à l’air externe. Il y a une masse qui limite le renouvellement complet de l’air de la caverne depuis l’air externe, d’où l’atmosphère différente. Le milieu dans lequel baignent les cellules des organismes pluricellulaires est soumis peu ou prou aux mêmes contraintes. Sa composition est différente de l’environnement. C’est ainsi que Claude Bernard a développé le concept de milieu intérieur.
Ce milieu intérieur permet aux organismes de vivre de manière plus indépendante de l’environnement. C’est ainsi d’ailleurs que des organismes peuvent vivre en milieu aérien, extrêmement desséchant. Les cellules échangent alors avec un fluide qui les entoure, appelé le liquide interstitiel. Or, il est nécessaire pour le bon fonctionnement des échanges et donc le bon fonctionnement des cellules que le milieu intérieur soit à une composition relativement constante. De faibles variations sont tolérables, mais les variations doivent rester de faible amplitude, selon le composé dont il s’agit.
C’est ainsi que Walter Cannon a développé le concept d’homéostasie du milieu intérieur, c’est-à-dire le maintien à un niveau constant des paramètres du milieu intérieur. Les besoins de l’homéostasie de ce milieu ont fait apparaître des organes spécialisés pour la constance du milieu. C’est ainsi que les poumons sont le lieu privilégié des échanges de gaz respiratoires ou encore les reins de la filtration du sang et l’élimination de certains déchets de l’organisme.
Les grandes fonctions physiologiques permettent donc l’homéostasie du milieu. Nous allons nous concentrer sur le rôle du cœur. L’homéostasie du milieu intérieur nécessite que les liquides internes de l’organisme passent par chaque organe spécialisé pour pouvoir en maintenir la constance. Ce fonctionnement est réalisé par des échanges en plusieurs étapes :
- dans un premier lieu, les cellules échangent avec le liquide interstitiel donc, comme nous l’avons vu
- le liquide interstitiel échange avec un autre liquide, le sang
- le sang passe par les différents organes et sa propre composition est maintenue constante
L’homéostasie nécessite donc l’existence d’un système de circulation du sang. Le sang circule sous pression grâce à l’existence d’une pompe, la pompe cardiaque. Le cœur assure ainsi la circulation sanguine par des cycles de contraction et permet ainsi la bonne réalisation des échanges. Dans cet article, nous chercherons à comprendre le fonctionnement cardiaque, et bien sûr des éléments de régulation de la fonction cardiaque qui asservissent le cœur à l’organisme et à son état de fonctionnement.
Ainsi, je vous propose d’étudier dans un premier lieu l’anatomie du cœur. Nous verrons ensuite les propriétés contractiles et l’émergence du rythme autonome du cœur.
L'anatomie cardiaque
Le cœur est un organe creux et relié assez logiquement au système circulatoire.
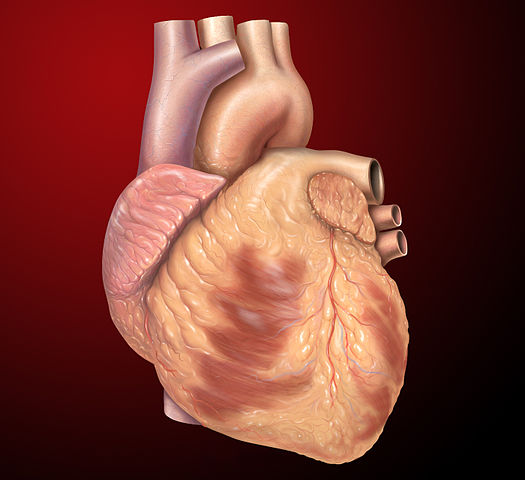
Sur cette image, on peut voir des artères et des veines ; on ne s’intéressera pas trop à ces vaisseaux dans la suite. Concentrons-nous d’abord sur le cœur. On peut voir deux grosses masses, une à droite et une à gauche, sachant que sur l’image la partie de droite semble bien plus développée. Il s’agit de deux unités qui sont séparées, mais fonctionnent de manière synchrone. On parle donc du cœur droit et du cœur gauche.
L’attribution des sens gauche et droit se fait dans le sens de l’organisme et non dans le sens où on l’observe. Puisqu’on observe le cœur par l’avant, ce qui est à droite sur l’image correspond à la gauche de l’organisme et réciproquement. Le cœur gauche est donc la masse de droite et le cœur droit, la masse de gauche.
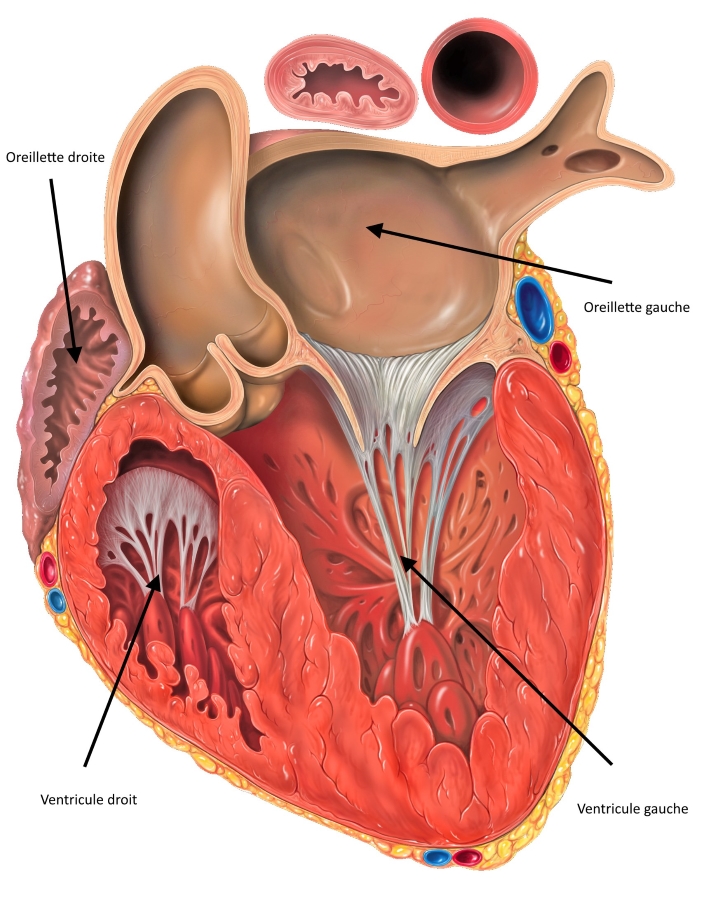
On peut voir sur ce schéma que le cœur droit comme le gauche se compose de deux parties : une oreillette et un ventricule. Le cœur se remplit au niveau de chaque oreillette (retour veineux au cœur), puis lors de la contraction l’oreillette expulse le sang vers le ventricule, qui éjecte ensuite le sang dans l’artère ressortant du cœur.
En anglais, ventricule se dit ventricle donc assez facile à comprendre, mais l’oreillette se dit atrium et cela conduit parfois à utiliser ce terme également en français. Comme la majorité de la littérature en français utilise le terme oreillette, c’est bien ce terme que nous utiliserons dans la suite du texte.
Chaque cavité est délimitée par une paroi composée d’une couche musculaire relativement importante (en rouge sur le schéma précédent) et d’une plus fine couche de revêtement de tissu conjonctif.
Le cœur est bien sûr irrigué lui-même par des vaisseaux sanguins (les artères et veines qu’on peut voir sur le schéma au niveau de la paroi). On parle des artères coronaires, à cause de leur forme dite en couronne autour du cœur (puisqu’elle suive la forme du cœur bien sûr).
Comme on peut le voir encore, la paroi musculaire du cœur gauche est bien plus développée que le droit :
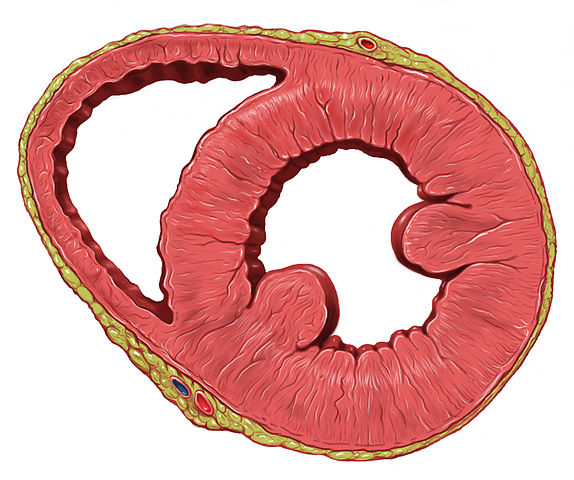
L’espace vide n’est bien évidemment pas vide, mais contient le sang. Le volume interne varie au cours du cycle cardiaque par l’alternance de remplissage et de constriction. Comme on peut s’y attendre, l’épaisseur plus importante du cœur gauche est liée à une force de contraction bien plus importante que celle du cœur droit.
Pour terminer cette partie, nous allons juste placer le cœur dans le corps.
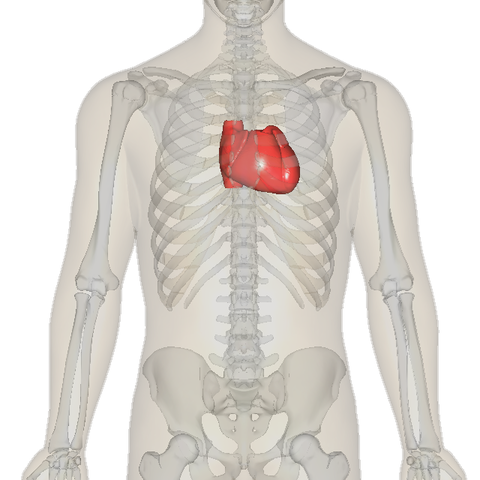
Ainsi, comme on peut le voir, le cœur se situe à proximité des poumons. On peut déjà relier cela à la légère atrophie du poumon gauche par rapport au droit (le cœur pointant à gauche)1.
Résumons donc ce qu’on a vu : le cœur se décompose en deux parties, droit et gauche. Chacun des deux cœurs est constitué d’une oreillette et d’un ventricule. Le sang arrive au cœur par les oreillettes et quitte le cœur par le ventricule. Il faudra donc s’intéresser à la circulation gauche et la circulation droite. Et il nous reste à faire fonctionner le cœur. On va continuer par parler des tissus et des cellules qui composent le cœur.
-
Sauf cas particuliers de situs inversus, où comme le nom le laisse penser, les organes sont inversés par rapport à la normale. ↩
Les cellules cardiaques
Comme tout organe, le cœur est constitué de plusieurs types cellulaires, dont les caractéristiques et la localisation à l’intérieur de l’organe sont importantes pour la fonction de l’organe. Il faut donc, pour le cœur :
- des cellules contractiles, capables d’éjecter le sang et de laisser le cœur se gonfler
- des cellules de soutien, qui vont développer un tissu conjonctif, résistant, qui va garantir une certaine rigidité aux endroits opportuns
- des cellules capables de développer le rythme intrinsèque de contraction
- des cellules capables de propager le signal de contraction
On va surtout parler des cellules contractiles et des cellules à activité rythmique ; les cellules de soutien ne sont finalement pas spécifiques du cœur, on parlera surtout du rôle du tissu conjonctif dans la contraction.
Les cellules contractiles
Ce sont des cellules de type musculaire (le cœur est avant tout un muscle !). Des cellules cependant différentes des cellules musculaires squelettiques, par leur organisation anatomique, leur structure, et même leur mécanisme de contraction. Cependant, elles en partagent certaines caractéristiques. On les nomme les cardiomyocytes (cardio pour le cœur, myo pour la contraction et cytes les cellules en grec). Ces cellules sont localisées dans la paroi, et en forment la partie musculaire contractile.

D’une part, elle sont bien évidemment contractiles, c’est-à-dire qu’elles peuvent raccourcir leur longueur. Les filaments à l’intérieur des cardiomyocytes (visibles sur le schéma précédent) sont impliqués dans ce raccourcissement. Les cardiomyocytes sont reliés les uns aux autres, donc le raccourcissement des cardiomyocytes, avec l’organisation vue précédemment, va diminuer le volume de la cavité cardiaque. Cela induit une hausse de la pression à l’intérieur de la cavité et c’est ainsi que les oreillettes et les ventricules expulsent le sang. Cette contraction est sous le contrôle d’un signal électrique.
D’autre part, les cardiomyocytes sont des cellules capables de propager les signaux électriques. Un cardiomyocyte reçoit un stimulus électrique, le propage sur l’ensemble de sa longueur, se contracte et propage le stimulus aux cardiomyocytes voisins. Les jonctions entre les cardiomyocytes sont permissives aux signaux électriques : on parle de synapse électrique. La propagation des stimulus électriques par ces cellules se faisant très rapidement, tous les cardiomyocytes d’une même cavité se contractent de façon quasiment synchrone.
Les quatre cavités ne sont cependant pas toutes synchrones, car les cardiomyocytes d’une même cavité sont isolés par du tissu conjonctif qui ne conduit pas les signaux électriques. Les oreillettes et les ventricules ne se contractent pas en même temps.
Des cellules de conduction électrique non contractiles
Les cardiomyocytes ne sont pas les seules cellules à être capables de propager un signal électrique. De nombreuses autres cellules le peuvent. Les neurones par exemple. Dans le cas du cœur, on va parler d’autres cellules encore, les cellules cardionectrices, qui forment différentes structures, parmi lesquelles par exemple le faisceau de His et le réseau de Purkinje.
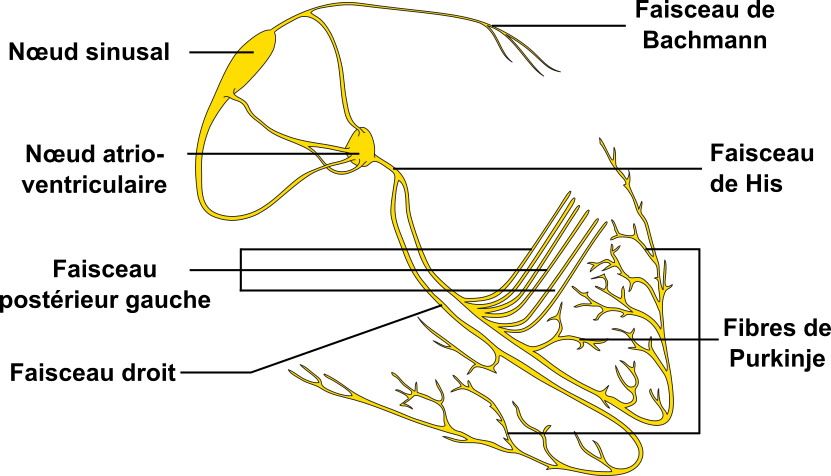
Ici, ces cellules ne servent qu’à la propagation du stimulus électrique (pas de contraction). Elles propagent les stimulus électriques mais moins vite que les cardiomyocytes. Cela va induire un retard dans la propgation et a une importance pour le cycle cardiaque, qu’on verra un peu plus loin.
Les cellules nodales
Ce sont les cellules qui composent les nœuds sinu-atrial (ou sinusal) et atrioventriculaire vus sur le schéma précédent ; pour plus de clarté sur leur localisation et leur interconnexion avec la partie musculaire, voici un schéma les replaçant dans le cœur.
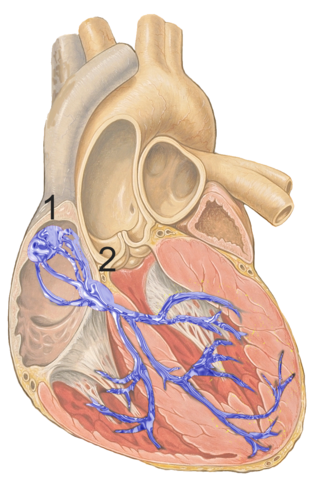
Ces cellules, en plus de pouvoir propager un signal électrique aux cellules suivantes, peuvent générer leurs propres stimulus électriques. Elles développent donc un rythme, qui entraînera l’ensemble du cœur. Ces cellules ont une activité dite pacemaker.
Les cellules nodales sont capables de générer leurs propres dépolarisations, contrairement aux cardiomyocytes et cardionectrices qui ne peuvent que propager mais pas initier le stimulus. Le cœur est donc capable de battre en absence de stimulus nerveux : si on réalise l’ablation du cœur expérimentalement, il continue de battre, malgré sa séparation du système nerveux de l’organisme.
Cela ne signifie pas que le cœur fonctionne indépendamment du système nerveux. Cela signifie simplement que le cœur n’a pas besoin de stimulus nerveux pour battre. Le système nerveux intervient dans son contrôle.
Ce sont ces dépolarisations, qui ensuite déclenchent les effets sur les cellules dépolarisées. Les cellules cardionectrices propagent les dépolarisations et les cardiomyocytes les propagent en même temps qu’ils se contractent.
Si on résume ce qu’on a vu, on a les supports de la contraction cardiaque avec les cellules musculaires cardiaques, les cellules de propagation des signaux électriques et on a vu que le cœur contient des cellules qui isolent les quatre cavités les unes des autres électriquement. On doit maintenant faire fonctionner le cœur pleinement, avec un cycle de contraction.
Le cycle cardiaque
Le cycle cardiaque comprend l’ensemble des événements d’une expulsion du sang du cœur (la systole) à la prochaine expulsion, d’où cette notion de cycle. On doit donc comprendre l’arrivée du sang aux oreillettes, l’éjection du sang dans le ventricule depuis l’oreillette et l’éjection depuis le ventricule dans l’artère. Voici, pour commencer, une animation de contraction cardiaque :
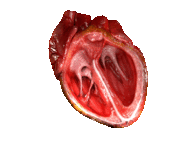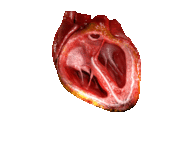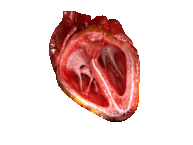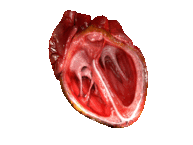
Comme on peut le voir, les contractions des ventricules suivent les contractions des oreillettes (dans cet ordre). Cela est permis par la vitesse de conduction plus faible du faisceau de His et du réseau de Purkinje : l’information de contraction passe obligatoirement par ces faisceaux (puisque les tissus conjonctifs ne conduisent pas l’électricité), et leur vitesse de conduction plus faible permet le décalage dans la contraction.
Les masses blanches, les valvules, sont du tissu conjonctif. Elles forment un pore qui ne s’ouvre que pour éjecter le sang dans un seul sens ; c’est un système anti-retour. On en retrouve entre les oreillettes et les ventricules, et entre les ventricules et les artères efférentes.
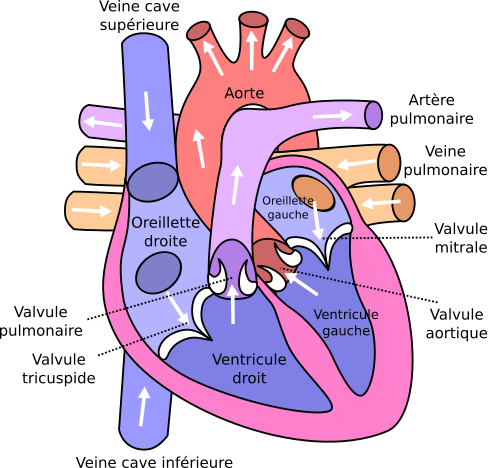
Pour illustrer le système anti-retour des valvules, étudions la valvule mitrale, du cœur gauche :
- lorsque l’oreillette gauche se contracte, la pression à l’intérieur de l’oreillette augmente et la valvule s’ouvre. Le sang passe de l’oreillette au ventricule.
- lorsque le ventricule se contracte, il y a un risque de retour sanguin vers l’oreillette. Or, l’augmentation de la pression ventriculaire exerce alors une force sur la valvule. Sa disposition est telle que les composantes de la valvule sont plaqués les uns contre les autres. Le ventricule n’est donc plus en communication avec l’oreillette : le sang ne peut que quitter le cœur par l’artère aortique, dont la valvule aortique est alors ouverte de la même façon que la valvule mitrale l’était par la contraction de l’oreillette à l’étape précédente.
Ainsi, on peut comprendre le trajet du sang. Le sang arrive au cœur droit par la veine cave, remplit l’oreillette droite qui se contracte et remplit le ventricule droit, puis part vers l’artère pulmonaire. Pour le cœur gauche, même principe, si ce n’est que le sang arrive par la veine pulmonaire et quitte le cœur par l’aorte.
Les signaux électriques de contraction vérifient le schéma de conduction suivant :
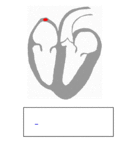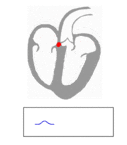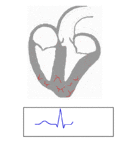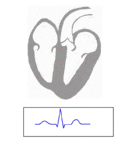
Une contraction ayant été faite, le cœur retourne à sa phase de remplissage (la diastole) et est prêt à initier un nouveau cycle de contraction. On peut suivre l’évolution de la pression intracardiaque et du volume du cœur au cours du cycle pour pleinement caractériser le fonctionnement cardiaque.
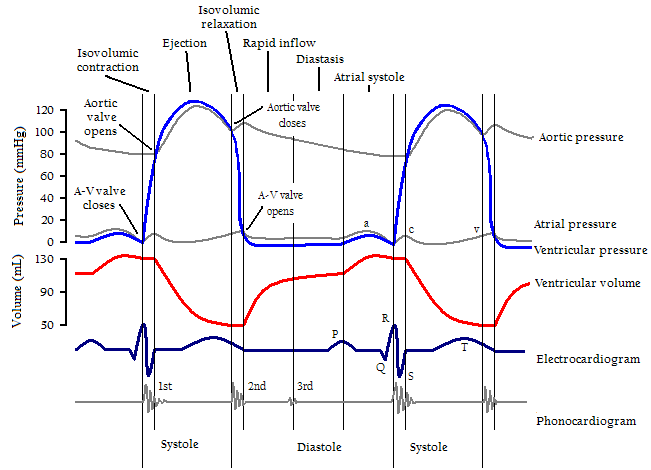
Le volume d’éjection correspond logiquement à la différence de volume du cœur entre la fin de remplissage (diastole) et l’éjection (systole). Tout le sang intraventriculaire n’est pas éjecté. On peut suivre la progression du cycle par les activités électriques (l’électrocardiogramme) ou par le bruit produit par l’ouverture et la fermeture des valvules (le phonocardiogramme).
Cela termine notre aperçu sur le cycle cardiaque. Nous avons essayé de faire fonctionner l’ensemble des tissus cardiaques pour expliquer sa contraction. Dans la partie suivante, on va essayer de relier le fonctionnement cardiaque au fonctionnement de l’organisme et donc changer d’échelle.
Le devenir du sang et les propriétés d'hémodynamique
Aspects généraux de la circulation
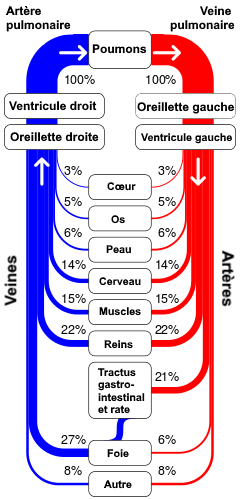
Comme nous n’avons cessé de le voir, le cœur est composé du cœur gauche et du cœur droit. Ces deux cœurs ont un fonctionnement synchrone mais sont séparés : ils transportent du sang qui n’a pas la même provenance depuis l’organisme et qui n’auront pas la même direction.
On ne verra pas trop ici les aspects circulatoires, nous allons nous contenter de décrire sommairement le devenir du sang. Les deux cœurs fonctionnent en série, l’un envoie le sang vers les poumons pour les échanges gazeux, l’autre reçoit ce sang fraîchement oxygéné et l’envoie vers l’ensemble de l’organisme.
Aspects d’hémodynamique
Nous avons vu que le cœur bat à un certain rythme et éjecte un certain volume à chaque cycle. À des temps courts, on peut supposer que les cycles vont avoir des valeurs à peu près constantes pour le volume d’éjection systolique et la fréquence cardiaque. On notera par la suite $FC$ la fréquence cardiaque (en coups par minute) et $V_{ES}$ le volume d’éjection systolique (mL).
Le cœur développe ainsi un mouvement du sang. La dynamique des fluides nous explique qu’on a alors affaire à un débit : une unité de volume qui arrive et quitte un point par unité de temps. Le débit sanguin correspond donc au débit cardiaque, $\dot Q$, développé par le cœur. Il s’agit donc du volume éjecté par le cœur par unité de temps.
À chaque cycle, le cœur expulse $V_{ES}$ sachant qu’il y a $FC$ coups par minute. Le débit sanguin, en mL/min, vaut alors
La fréquence et le volume d’éjection systolique sont deux paramètres sur lesquels il peut y avoir des régulations en vue de la régulation du débit cardiaque. Plus le cœur bat vite et plus il éjecte de sang par unité de temps et plus le débit sanguin est important.
Ce n’est pas tout à fait le débit sanguin qui est régulé mais la pression artérielle, à laquelle il est lié.
La fréquence moyenne est d’environ 70 coups par minute et le volume d’éjection moyen est de 70 mL. On retrouve alors un débit sanguin moyen de 5L/min. Naturellement, cela dépend des individus et de l’état de fonctionnement de l’organisme (en cas d’effort physique intense par exemple).
Par ailleurs, la dynamique des fluides a été très étudiée en physique. On va chercher à exploiter certaines propriétés pour expliquer certaines propriétés du système circulatoire.
Les propriétés de dynamique des fluides supposent que le liquide circule de façon continue ; or le cœur fonctionne par à-coups. Il s’agit du travail des grosses artères de sortie du cœur de lisser le débit discontinu en débit continu. Par la suite, on fera comme si ce lissage du débit a été effectué.

Sous certaines conditions qu’on supposera applicable pour le sang et le système circulatoire, les différents paramètres mentionnés sur le schéma sont reliés par la loi de Poiseuille appliquée à un cylindre :
Ce qu’on réécrit sous la forme de la loi de Darcy appliquée au système circulatoire, entre la sortie et l’entrée du cœur (l’ensemble du système circulatoire est assimilé à un simple cylindre) :
La valeur de la résistance périphérique est déterminée par le système circulatoire, la fréquence cardiaque et le volume d’éjection par le cœur. Ce n’est pas le débit sanguin qui est contrôlé mais la pression artérielle. En cas de perturbation de la pression, le corps répond par des adaptations vasculaires et cardiaques.
Ces propriétés d’hémodynamique sont importantes pour expliquer l’écoulement du sang, c’est pourquoi j’en touche quelques mots mais nous allons refermer la parenthèse ici. On retiendra qu’effectivement la pression est l’élément moteur de l’écoulement du sang et que la pression est relié au débit sanguin, ce qui explique que la régulation de l’un régule l’autre.
Dans la partie qui suit, on va s’intéresser à quelques éléments de régulation du fonctionnement cardiaque.
Régulation cardiaque
Régulation autonome et réflexe de Starling
Le cœur est capable de réguler de son propre chef sa contraction. Ainsi, sous l’influence du retour sanguin dans l’oreillette, le cœur peut varier l’intensité de sa contraction. Cela a été décrit par Starling et ce réflexe autonome porte donc son nom :
Le cœur répond à une hausse du retour veineux par une hausse de sa force de contraction, ce qui accroît le volume d’éjection systolique $V_{ES}$.
Le support de ce réflexe est l’extension des cardiomyocytes par le retour veineux ; ces cellules répondent à la contrainte de déformation par cette hausse de puissance de contraction. Ainsi, dans un premier temps on a d’abord l’oreillette qui se contracte plus violemment, qui dilate alors le ventricule davantage et le ventricule augmente à son tour sa force de contraction.
Par ailleurs, puisque le support du réflexe de Starling est la distension des cardiomyocyte, chacun des deux cœurs fonctionne indépendamment de l’autre quant à ce réflexe et cela garantit le bon fonctionnement du cœur. Si un des cœurs délivre un débit plus important que l’autre, il y aura accumulation de sang dans la circulation pulmonaire ou dans la circulation générale, par rapport à l’autre. Prenons le cas où le cœur droit a délivré un débit plus faible que le gauche pendant un certain temps. Alors, le cœur gauche reçoit moins de sang et en conséquence, il se contracte moins : le volume éjecté par le cœur gauche diminue, sans impacter celui du cœur droit. Cela compense alors le déséquilibre initial entre les deux cœurs.
Le réflexe de Starling permet un équilibre du fonctionnement cardiaque et quelques éléments de régulation de la pression artérielle. Si la pression artérielle augmente, le plasma (la partie liquide du sang) passe dans les tissus par filtration au niveau des vaisseaux (imaginez un système poreux dans lequel on fait passer un liquide : il passe à travers les pores et moins de liquide ressort de l’autre côté du tube) ; donc le retour veineux au cœur est diminué et le cœur éjecte moins de sang, ce qui tend à faire diminuer la pression artérielle.
Ce réflexe permet aussi d’expliquer la baisse de la fréquence cardiaque chez les sportifs entraînés : en réponse à l’entraînement, il y a une hausse de la quantité de sang de l’organisme (hypervolémie), le retour veineux est augmenté donc le cœur se contracte plus. Pour ramener la pression à une valeur normale, l’organisme diminue la fréquence cardiaque.
Régulation nerveuse
Le cœur est innervé par les deux branches du système nerveux autonome : le système nerveux sympathique et parasympathique. Le système parasympathique tend à diminuer l’activité cardiaque, le sympathique à l’accroître.
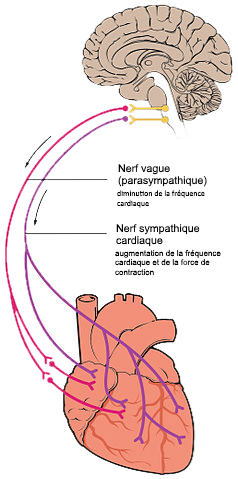
Les deux systèmes se distinguent par leurs organisations (qu’on ne détaillera pas ici) et par le neurotransmetteur qu’ils émettent. La transmission d’information par le système nerveux se fait par l’émission dans l’espace intercellulaire d’un neurotransmetteur, qui agit sur les cellules avoisinantes via leurs récepteurs. La fixation d’un neurotransmetteur déclenche une réponse cellulaire, différente selon le récepteur activé. C’est ainsi, grâce à l’utilisation de deux neurotransmetteurs différents, que de mêmes cellules peuvent recevoir et réagir à des signaux antagonistes.
Le système parasympathique utilise l’acétylcholine, le sympathique la noradrénaline. L’acétylcholine diminue la vitesse d’émission des stimulus électriques par les nœuds (effet chronotrope négatif). La noradrénaline accroît la vitesse d’émission de ces stimulus (effet chronotrope positif) mais également la force de contraction des cardiomyocytes et donc, le volume d’éjection systolique (effet inotrope positif).
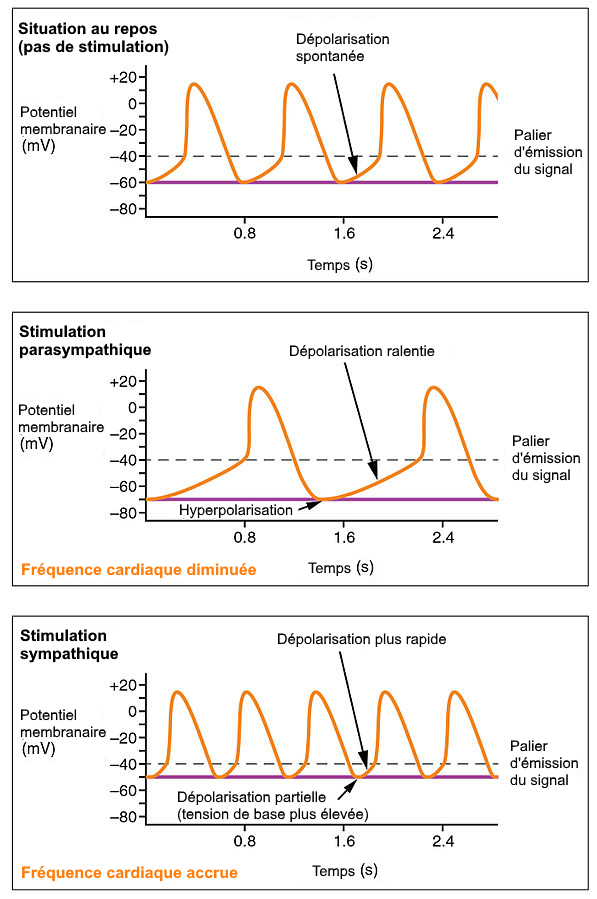
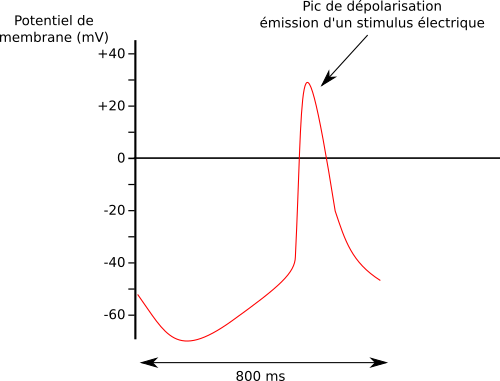
En absence de stimulus nerveux, la tension basale est vers -60mV ; la pente de dépolarisation a un coefficient basal qu’on qualifiera de moyen. Avec la stimulation parasympathique, la tension basale est plus basse, donc il faut un temps plus long pour une dépolarisation. Par ailleurs, le coefficient de dépolarisation est abaissé donc la dépolarisation est plus lente. La fréquence d’émission des signaux électriques est donc plus faible.
A contrario, la stimulation sympathique tend à réhausser la tension basale des cellules nodales (vers -50mV sur l’image), donc il faut moins de temps pour dépolariser. Et le coefficient de la pente de dépolarisation est accru donc la dépolarisation est plus rapide. La fréquence cardiaque est accrue. Il ne faut pas oublier par ailleurs que la stimulation sympathique accroît également le volume d’éjection systolique.
Ainsi, le système nerveux parasympathique permet une diminution de la pression artérielle, le sympathique une augmentation.
Régulation hormonale
Des hormones circulantes peuvent également agir sur le cœur. L’adrénaline est libérée en situation de stress et a exactement la même action que la noradrénaline sur le cœur. Elle fait donc redondance avec la noradrénaline et assure une bonne stimulation cardiaque, en cas de stress (émotionnel ou physique, tel qu’une hémorragie par exemple).
Exemple fonctionnel : le baroréflexe
La régulation de la pression artérielle est essentielle au fonctionnement de l’organisme. Un défaut de pression peut entraîner un manque d’irrigation sanguine des tissus, tandis qu’une trop forte pression peut entraîner un éclatement des vaisseaux et provoquer une hémorragie. En conséquence, il existe de nombreux systèmes de régulation de la pression artérielle. On a pu apercevoir quelques éléments de régulation, de manière diffuse au cours de cet article.
Comme on l’a vu avec la loi de Darcy $\Delta P = FC \times V_{ES} \times R_{périphérique}$, la régulation de la pression peut faire intervenir le cœur et les vaisseaux sanguins. On s’intéressera toujours uniquement aux effets sur le cœur.
Il existe des moyens de régulation à court, moyen et long-terme. Le baroréflexe est un exemple assez simple de réflexe autonome de régulation de la pression artérielle et agit à court-terme. Une variation rapide de la pression artérielle engendre une réponse rapide du système nerveux qui contre-balance cette variation. Il intervient typiquement lorsqu’on passe de la position couchée à la position debout rapidement : le changement de position nécessite de lutter contre la gravité. La pression artérielle chute légèrement et il faut une compensation rapide. Elle est assurée par le baroréflexe.
Prenons un cas où la pression artérielle a augmentée :
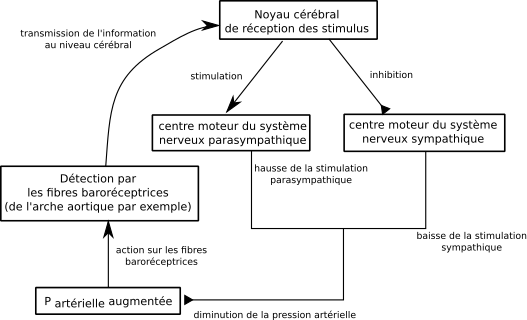
Le baroréflexe entraîne également des adaptations vasculaires, ce qui fait que l’ensemble des éléments intervenants dans la pression artérielle est impliqué dans la correction de la variation de pression. Les effets anatomiques permettent de corriger la perturbation initiale, on parle de boucles de rétroncontrôle négatif.
Le cœur est ainsi asservi au fonctionnement de l’organisme et assure à la fois la circulation sanguine et la régulation de la pression artérielle.
Dans cet article, nous avons essayé de relier la structure du cœur à son fonctionnement et à comprendre l’intégration du fonctionnement cardiaque à l’organisme. Nous avons ainsi vu les cellules cardiaques, leurs propriétés globales et vu comment leurs interconnexions fonctionnalisent l’ensemble.
Nous avons vu ensuite que des propriétés physiologiques émergent du fonctionnement de l’organe avec la pression artérielle et les propriétés dites d’hémodynamique. Le cœur étant asservi à l’organisme et toujours dans une optique d’homéostasie du fonctionnement de l’organisme, la pression artérielle est étroitement régulée.
J’espère que ces paragraphes vous auront intéressés et que le fonctionnement cardiaque est quelque peu démêlé ; cet article ne se voulait pas exhaustif mais plutôt une introduction à la physiologie cardiaque. À ce propos, concernant l’approfondissement du sujet, cet article a été écrit en s’inspirant de ces deux livres :
- Medical Physiology, Boron & Boulpaep, Elsevier, 2016, 3è édition
- An Introduction to Cardiovascular Physiology, Levick, Hodder Arnold, 2009, 5è édition
L’ensemble des illustrations proviennent de Wikipédia et sont libres de droits (licences CC0, CC BY ou CC BY-SA).
Articles de wikipédia depuis lesquls sont tirés les images :
- Cœur (en et fr)
- Cellule musculaire cardiaque (en)
- Faisceau de His (fr et en)
- Nœud sinusal (en)
- Système circulatoire (en)
- Cycle cardiaque (en)
- Fréquence cardiaque (en)


